In spite of the overwhelming evidence against it, scientific creationism is a growing movement (Numbers, 1982). Numerous authors (for example, Kitcher, 1982; Futuyma, 1982; Godfrey, 1983, 1984; Montagu, 1984) have argued that scientific creationists use questionable tactics to achieve this success. Creationists have been accused of distorting the evidence, misrepresenting evolutionary scientists, and being ignorant of both evolutionary biology and the philosophy of science.
In answering creationists, it is understandable that we should want to use the latest facts that research has put at our disposal. Yet, curiously, in doing so we may be playing into their hands. This sounds very strange, so let me explain.
What we would be doing is admitting to the tacit assumption that debate simply hinges on what the facts are. This would be a mistake, I think. As important as the new facts are, they are not needed to show that evolution is superior to scientific creationism. There was enough evidence in Darwin's day. What is needed is clear thinking. Darwin displayed this. I will show that creationists don't.
Indeed, one of the tactics of creationists is to dazzle with carefully chosen facts and to couch their arguments in such highly technical language that the fallacies in their thinking are often obscured. My aim here is to show that creationists make apparent headway with their arguments because they conflate three distinct types of evidence. To demonstrate their fuzzy thinking, it will be helpful to consider an illustration.
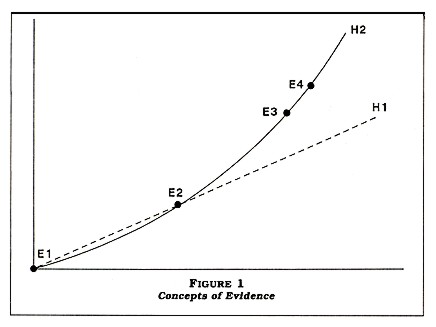
In Figure 1, facts are represented by dots on a graph (see page 16). Hypotheses are represented by curves. If we were given only E1 and E2, there would be no way of knowing which is the better hypothesis, H1 or H2. Since E1 and E2 are common to both hypotheses, let's call this evidence common evidence. To determine which is the better hypothesis, we need a different kind of evidence. This second type we will call discriminatory evidence (E3 and E4). Given this new evidence, we can easily see that H2 is superior to H1. The discriminatory evidence (E3 and E4) has decided the issue and is therefore an important and distinct type of evidence. Anderson (1987) defines these two types of evidence as follows: common evidence is that kind of evidence which is equally favorable to two or more hypotheses, and discriminatory evidence is that kind of evidence which tends to favor one hypothesis over its rival.
Have we proved that H2 is the correct hypothesis? No, not in any ultimate sense. But we have shown that H2 is the better of the two alternatives. To create the impression that H1 is equally acceptable as a scientific hypothesis, one might use the following tactics.
Tactic One
One could belabor the point that there exists evidence which is actually common to both alternatives and which precisely fits one's theory. Of course, it is of no consequence that E1 and E2 precisely fit H1; it is the remaining evidence which is critical. This is a species of a well-known fallacy called special pleading. Creationists commit this fallacy, as the following examples will show.
The first example of creationist special pleading is their use of apparent design. Apparent design is evidence which is common to both evolution and creation. Evolution accounts for design through natural selection acting on heritable variation.
Scientific creationism claims that design is evidence of special divine intervention. As both alternatives account for design, this is common evidence, and we cannot rely on it. Yet, this is exactly what creationists do. For instance, Coppedge writes: "There is no way chance mutations could have brought about the development of feathers. Each is a masterpiece of engineering" (1977, p. 215).
Of course, according to evolutionary theory, it is not chance mutations that bring about development but natural selection (a powerful anti-chance force) acting upon variability. Even this is a very simplified picture. That apparent design can be accounted for by evolutionary theory is very well attested to. The dark form of the moth Biston betularia is camouflage against the dark polluted bark of trees. The "camouflage design" is apparent and is merely a result of selection instead of the pale form. Therefore, as Cracraft points out: "Creationists should not invoke common design because . . . statements about functional design . . . are consistent with evolution as well" (1983, p. 176).
Similarity in living organisms is another example of common evidence used in creationist special pleading. For evolution, this similarity suggests common ancestry. For scientific creationism, this similarity suggests a common blueprint. Because both alternatives can account for similarity, we cannot rely on this evidence. Creationists frequently do. For example, Wilder-Smith writes: "Darwinism postulates that similarities in living organisms predicate common ancestry" and adds that this similarity might "be better explained on the basis of a common plan or planner behind them" (1974, pp. 52-53).
A third example of common evidence used in creationist special pleading is sterility in hybrids. According to evolution, sterility is a byproduct of constitutional differences between hybrids. Creationists can also explain sterility, as Darwin notes: "The view generally held by naturalists is that species, when intercrossed, have been specially endowed with the quality of sterility, in order to prevent the confusion of organic forms" (1859, p. 264). Since both alternatives can explain sterility, we should not rely on this line of evidence, as creationists did in Darwin's day.
Tactic Two
Let's return now to our illustration. We noted that H2 is the better hypothesis. The second tactic which could be used to create the impression that H1 is an acceptable scientific hypothesis is to gloss over or play down or try to explain away the discriminatory evidence. Once more, we find creationists doing this.
One example of discriminatory evidence is the imperfection in living things. If living things were specially created, we would expect them to be perfectly designed, and they are not. Whereas Darwin focused on the lack of fit between organism and environment (1859, pp. 216-217), creationists are revealingly silent about such imperfections in living things—for example, the webbed feet of upland geese, the absence of webbed feet in subaquatic thrushes, and the makeshift semi-opposable thumb of the panda (Gould, 1980, pp. 20-23). Coppedge (1973) and Frair and Davis (1983), for instance, completely ignore the imperfections in living things.
Another example of discriminatory evidence is the fact that the similarities in organisms form a hierarchical pattern. Creationists cannot account for this pattern, but it is easily explained through evolutionary descent, as Cracraft has pointed out:
The hierarchical pattern produced by the shared similarities observed among organisms is predicted by a hypothesis of evolutionary descent with modification but not by an assumption of special creation.
[1983, p. 172]
To my knowledge they [scientific creationists] have never denied the existence of a hierarchy of similarity, nor have they provided a credible, scientific explanation as an alternative to descent with modification.
[1983, p. 177]
A third example of discriminatory evidence is that the sterility of organisms is in degrees. Darwin states it well: "We see the truth of this conclusion" (that sterility is incidental on constitutional differences rather than a special endowment) "in the vast difference in the result, when the same two species are crossed reciprocally . . ." (1859, p. 436). Darwin did not invoke sterility as such in his defense of descent by modification; he realized that it was common evidence. Instead, he appealed to the fact that sterility between organisms is in degrees. Under special creation, one would expect absolute sterility between species, irrespective of organism and the direction of the cross.
Creationists try to explain away evidence that discriminates against their position by using ad hoc modifications of their hypothesis. There is the undisputed fact of the sequence in the fossil record of increasing numbers of more complex organisms. Creationists try to get around this evidence by positing a global flood. Whitcomb and Morris give three sorting mechanisms purportedly explaining the fossil sequence (1961, pp. 273-274), all of which involve flood geology. One is hydrodynamic sorting—the idea that denser and more streamlined objects would have descended more rapidly and should be found in the lowest rock strata. Another is ecological sorting—the idea that the inhabitants of low regions would be overcome first by the flood waters and would be in the lowest strata. The third mechanism they propose is anatomical sorting—the idea that the least mobile animals would have succumbed first to the ravages of the flood and would be in the lowest strata.
The problem with these three hypotheses is that they are mutually incompatible and false. Gould (1983) points out that the fossil record contradicts them. Whales, which are streamlined and marine, are found only in the upper strata, contradicting both the first and the second explanations. Sloths, which are not known for their mobility, are also found only in the highest strata, thus contradicting the third explanation.
Tactic Three
Returning once again to our original illustration, let's look at one more tactic that someone could use to give the impression that H1 is an acceptable scientific theory.
The "straight-liner" could claim that the incompleteness of the data for H2 constitutes evidence for H1. This claim makes at least three mistakes. First, it assumes that H2 and H1 are the only two alternatives. This is a fallacy in itself and goes by the name of the faulty dilemma. Second, the demands of the straight-liner are unreasonable. The straight-liner gives the impression of being very rigorous in requiring that the supporter of the exponential curve fill in the gaps. But it is easy to see the inconsistency. With regard to completeness of evidence, H1 is worse off. The principle violated is that hypotheses should be compared over equivalent domains. For the given set of coordinates, H2 is superior to H1. Third, the straight-liner claims that the issue is the amount of evidence when, in fact, it is the kind of evidence that is critical. Very little discriminatory evidence is needed to favor one hypothesis over a rival. If E1 through E4 are the only evidence we have, then E3 and E4 are all that is needed to tip the scales against H1.
Incidentally, we can see here why the straight-liner's approach might have a certain appeal. It is well known that creationists take advantage of a popular misconception that real science is pure fact (Kitcher, 1982). Since evolution is just a theory, they claim that it is not really science.
This illustration helps us to see that the straight-liner's problem is not a purely empirical one. Let us imagine that additional evidence E5 through E8 is found. Does this evidence compel acceptance of H2 over H1? Not to the straight-liner, who could point out that the lines drawn between E4 and E5, E5 and E6, and so forth, are interpolations and not facts. Therefore, H2 is not really factual and H1 and H2 are both unscientific. The straight-liner might say that it is quite in order to believe H1. No amount of evidence will convince such a person. Since the curve H2 is infinitely divisible, it can never consist of pure fact. The straight-liner's error is in holding to a discredited view of the nature of science. Popper has shown that universal statements (the bricks of scientific theories) can never be reduced to pure fact (1972, p. 36).
Creationists argue very much like the straight-liner. They start by claiming that the only two options are evolution and special creation (for example, Enoch, 1968, p. 105; Frair and Davis, 1983, p. 55). Then they say that evolution predicts a finely graded series of transitional forms instead of the gaps we find in the fossil record and that, therefore, the facts speak against evolution and for special creation. This argument is not uncommon:
It may therefore be asserted that the facts of paleontology, namely, the sudden appearance of all the invertebrates with their distinct specialization in the cambrian era, and the complete absence of undisputed fossils in the precambrian rocks, and the absence of intermediate forms linking the basic kinds of animals and plants, argue more for instantaneous creation than for evolution.
[Enoch, 1968, p. 55]
One example of the scientific evidence for creation is the sudden appearance of complex fossilized life in the fossil record, and the systematic gaps between fossilized kinds in that record. The most rational inference from this evidence is seemingly is that life was created and did not evolve.
[Gish, Bliss, and Bird, 1981]
(see also Frair and Davis, 1983, p. 56; Chittick, 1984, pp. 64-65; Gish, 1985).
Scientists have often responded to such claims by pointing out that creationists are ignorant of certain basic facts: that despite the imperfections in the fossil record (Raup, 1983, p. 158), there are Precambrian fossils (Godfrey, 1983, p. 198); that the emergence of life was only sudden on a geological time scale; and that there are transitional forms (Godfrey, 1983, pp. 198-202; Cuffey, 1984, p. 257; Burke, 1985).
I wish to emphasize that there are additional philosophical problems with the creationist arguments. To treat the problem as purely empirical is to fall into a creationist trap. They create a superficial impression that theirs is the truly empirical approach, when no amount of evidence will satisfy them. By setting up an impossible standard and putting the onus on the evolutionary scientist to meet this standard, they put the evolutionary scientist on the defensive. Our response to the creationist should not be just to display the evidence for evolution but also to expose their faulty philosophy of science.
The creationist mistake is that he or she has conflated negative and discriminatory evidence. Because it is unreasonable to demand complete evidence, negative evidence does not count against a theory. That a significant part of the creationist problem is a philosophical one can be seen from the way they treat transitional forms.
Archaeopteryx is a classic transitional form, showing both reptilian and avian features. It is interesting to note that the responses of creationists to this form are quite divergent. One objection is that Archaeopteryx is not a true intermediate. Creationist Baker says, "If archaeopteryx has got a fully reptilian skeleton, then it has certainly got feathers that are fully bird-like. There is no suggestion of something at the half-way stage between a reptilian scale and a bird feather, which is what an intermediate creature would be expected to possess" (1976, p. 12). This sort of objection was addressed by Darwin (1859), of course, so we won't let it detain us here.
For our purposes, a second objection is most interesting. Cousins claims that Archaeopteryx is not a true missing link but, rather, an aberrant bird (1971, pp. 89-99). We can dispute this interpretation, of course, but the tactic of interest to us here is that, when a major gradation is found, the absence of finer gradations is taken to be evidence against evolution. If such finer gradations are found, the creationist can then point to the absence of even finer gradations, and so on. Any evidence can always be explained away because negative evidence will always be available. The creationist's mistake is in giving negative evidence the status of discriminatory evidence.
Conclusion
Complete evidence is not needed to decide between rival hypotheses—only discriminatory evidence, and not much of it. There was enough in Darwin's day to decide between evolution and creationism. In order to show that evolution is superior to the type of creationism promulgated by Morris and Gish, one need only show that, as one goes further back in the fossil record, the fossil forms become progressively different from the forms existing today. That is all. One does not need to show a finely graded series with innumerable links. Special creation, in contrast, does not predict this order., To do so, all forms would be uniformly distributed throughout the fossil record—and they are not.
The superiority of evolutionary theory is evident in spite of an imperfect body of evidence for it. Darwin realized, at least to some extent, that discriminatory evidence, rather than negative evidence, is needed to adjudicate between scientific hypotheses. Ghiselin says that Darwin rejected particular explanatory hypotheses in biogeography "not because some evidence which might support them is not available, but because the facts are better explained by some contrary hypothesis" (1969, p. 42). Furthermore, we find that Darwin admitted that the fossil record was incomplete and yet was able to see that it was sufficient to favor descent by modification over independent creation (1859, pp. 291-316). He wrote:
Let us now look to the mutual affinities of extinct and living species. They fall into one grand natural system; and this fact is at once explained on the principle of descent. The more ancient any form is, the more, as a general rule, it differs from living forms.
[p. 330]
The main point is this: we do not need more , new evidence to show that evolution is superior to scientific creationism. The evidence we need has been around for a long time and is even accepted by creationists. They do not deny, for instance, that the fossil record shows a sequence of simple to complex forms. What we do need is clear thinking. Whereas scientific creationists fail to distinguish between common and discriminatory evidence and between negative and discriminatory evidence, Darwin succeeded. Others have come to similar conclusions (see, for example, Kitcher, 1982, pp. 30-54; Futuyma, 1982, pp. 175-196). In the following statement, it is evident that Futuyma comes to the same sort of conclusion as mine, although he does not give the types of evidence appropriate names. He writes:
In actuality, almost all creationist literature simply consists of attacks on evolution, rather than positive evidence for creation. To the creationists any [negative] evidence against evolutionary theory apparently constituted [discriminatory] evidence in favour of creation."
[1982, p. 176]