Especially since the publication of Henry Morris and John Whitcomb’s book The Genesis Flood (1961), young-earth creationists have claimed a scientific basis for their view that a historic, worldwide flood shaped the major geophysical features of the earth. A key problem for "flood geologists" is the order and sequence of fossil remains in the geological record. In order to construe the bulk of the geologic column as a result of a single global deluge, flood geologists must replace the current scientific explanation of the fossil record as a result of preservation of series of contemporaneous ecological associations of organisms with one that is consistent with the aftermath of a major geologic catastrophe.
Modern taphonomic studies clearly distinguish the patterns of deposition of organic remains after severe modern flooding from those produced by other processes. The fact that the bulk of the fossil record does not show evidence of the sudden, catastrophic deposition expected from a massive flood means that most of the fossiliferous strata were laid down by some other geologic process. Furthermore, contravening the expectations from a large flood, the fossil record records a succession in the history of life consistent with a theory that is based on the emergence of new species via descent with modification from common ancestors.
A critical requirement of the flood model is to provide a reasonable explanation for this observed stratigraphic succession of both flora and fauna — a universal feature of the fossil record. In an attempt to solve this problem, flood model advocates quickly seized upon the fact that ecosystems naturally vary; as one travels from the seashore to the highlands or from the equator to the poles, the characteristic plants and animals that make up ecological communities will also vary. Invoking this well-documented observation, flood model advocates applied the term "ecological zonation" (EZ) to the hypothesis they developed to account for the observed stratigraphic succession of organisms.
In brief, EZ postulates that, as the water of the Flood rose, organisms were buried according to the ecological zones in which they lived. Thus, according to flood model proponents, marine organisms would be buried first, as sediments derived from the breakdown of rocks that formed the land flowed into the continental shelf and farther into the ocean basins. The next layer would be organisms from near-shore terrestrial environments, and as the water continued to rise, organisms from higher elevations would finally fall victim, or so the explanation goes. Thus, EZ is supposed to account for the well-documented biostratigraphic succession from marine organisms to amphibians and on to organisms fully adapted to terrestrial environments. The development of EZ in the first place is a recognition by flood geologists that the fossil record does not look at all like the outcome of a large-scale flood.
Naturally, flood geologists have overlooked a few facts that demonstrate that EZ is untenable. I will review two significant flaws in the EZ hypothesis that also expose the fundamental explanatory failure of the flood model: its inability to explain the distribution of organisms (the biostratigraphic order) and the pattern of the layers of rocks (the lithostratigraphic order) of the geologic column.
EZ doesn’t do it
The first argument rests upon the observations of the geological record for North America, as summarized in the Geological Society of America’s Decade of North American Geology (DNAG is a series of books about the physiographic geology of North America issued through the 1980s in commemoration of the 100th anniversary of GSA, including an authoritative timescale as of 1986). These observations document that in the past, during most of the early Paleozoic Era, most of North America was covered by broad shallow (epeiric) seas. Indeed, geological atlases show that most of the North American continental interior and continental shield (craton) was covered by these shallow seas and was of very low relief — a relatively flat seabed, with no major valleys or mountains. By contrast, the area known as the Canadian Shield — a large region of Proterozoic rock with little in the way of more recent fossil or sedimentary deposits — was apparently not covered by these shallow seas.
Thus, in North America the Cambrian Period (see Table 1), for example, is dominated by strata bearing organisms of marine origin. Similarly, the Ordovician and Silurian periods are also composed almost exclusively of marine deposits. Later, the Devonian and Carboniferous periods are also composed of large amounts of marine strata, except for deposits near the Appalachians and part of what would become the Rocky Mountains. This is because a series of continental collisions that occurred during that time period formed the Appalachians, proto-Rockies, and several other areas of significant relief.
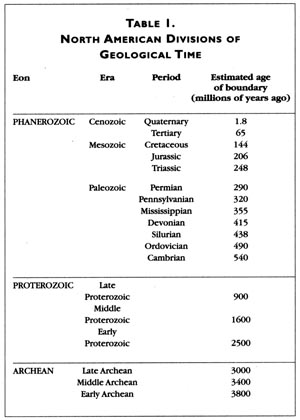
Except for the Canadian Shield, the oldest North American deposits in undisturbed strata always contain marine organisms; terrestrial organisms only occur in more recent strata. Evolutionary theory accounts for this by the successive diversification of descendants that colonized the land from ancestors who were solely or chiefly living in the water. There are no undisturbed strata in which organisms from, say, the Phanerozoic are mixed with or appear in strata older than those of the Proterozoic.
This observation is extremely damaging for young-earth creationists promoting EZ during Noah’s Flood to explain the distribution of rock layers and organisms in the geologic record. Even the work of John Woodmorappe (who is also known as Jan Peczkis; 1999) that attempts to discredit the geologic column as an artifice built up by loose deductive reasoning inadvertently confirms that the Canadian Shield constituted dry land during the Phanerozoic. It also confirms that the rest of North America was covered with water during the purported "pre-Flood" period. This means that the terrestrial organisms in the sediments throughout North America had to have been transported thousands of kilometers from their habitats on the Canadian Shield to their final deposition locations. To accomplish this, somehow terrestrial vertebrates and plants had to have been uprooted from their original locations, floated into their present positions, and then settled out of the water carrying them to be deposited in a way that preserves the major components of the biocenosis (a natural assemblage of organisms or a living community).
But this raises other serious problems for EZ — and puts this model at odds with another creationist "explanation" for the geologic column, hydrodynamic sorting (HS). Proponents of HS argue that organisms that can float, swim, or even outrun flood waters will be found higher in the geologic column that those that cannot. How is it possible, then, for organisms with such divergent hydrodynamic properties first to be transported together long distances then later to settle to the bottom and be deposited in a manner that preserves local ecological communities? EZ fails to explain how (younger) rocks of the Mesozoic and the Cenozoic eras have come to have fossil terrestrial organisms situated in their proper ecological context, rather than as mixed assemblages of fossil organisms whose ecological relationships with other organisms have been disrupted by the violent waters of the Flood.
Furthermore, how can transport and burial of nearly all terrestrial flora and fauna come to mimic the order that evolutionary theory would require (for example, assemblages of marine organisms in earlier deposits, with near-shore amphibians, then reptiles, and then birds and mammals, each associated with its own particular flora in successively later deposits)? Flood model advocates embrace EZ to explain this succession, but with almost all of North America covered by epeiric seas prior to the onset of the Flood, as even Woodmorappe’s study (1999) shows, they are forced to accept that the only place where EZ could operate as envisioned for terrestrial organisms is in the area of the Canadian Shield, from where the organisms would be transported by the Flood waters over the rest of North America.
One of the biggest challenges for EZ is the "dinosaur freeway" documented by Martin Lockley (Lockley and others 1992). For example, the Caririchnium ichnofacies (a rock unit characterized by a distinctive suite of trace fossils) is a megatrackway consisting of the footprints of sauropods and theropods (plant-eating and meat-eating dinosaurs, respectively) can be found over an area of 80 000 square kilometers. Of course, the Caririchnium ichnofacies and numerous other similar ichnofacies were formed by the actions of many dinosaurs walking on dry land.
But on the flood model, the terrestrial animals responsible for these tracks must have been transported by water from terrestrial habitats far removed from the areas where the trackways were produced. To make that journey, these animals had to survive long-distance water transport, touch down over a wide area on the North American continent, and somehow make tracks on the seabed before they all perished in the Flood. Only wildly ad hoc hypotheses seem capable of explaining the survival of the organisms until they reach the site, or the creation of these tracks, especially given that all of these areas were already under the waters of the epeiric seas prior to the arrival of the organisms (see, for example, "Bibliolatry revisited" by Wilfred Elders, p 33). Of course, the standard — and most rational — conclusion is that the area that was a seabed in the Paleozoic was a terrestrial habitat much later and that the tracks were made by the animals that lived on the land then.
There are more problems for the transport model when we consider the presence of paleosols (ancient soil horizons preserved in the fossil record). In these cases, the soils are arranged in distinguishable layers, clearly an in situ feature. In addition, many paleosols have preserved traces of root systems, including even fine-scale features such as rootlets. Examples of paleosols from the Mesozoic include those of the Triassic Delores Formation documented by Blodgett (1988) and various Cretaceous paleosols studied by Sigleo (1988), to name but a few. If these soils had to endure transport over thousands of kilometers from the Canadian Shield, how did they re-assemble as distinct layers after being mixed up by the violent action of flood waters — including re-assembly of root systems?
So it is easy to see that flood geology is at odds with most of the observations of North American geology. EZ is one model that has been proposed to reconcile these observations with the flood model, but EZ only works if inappropriate ad hoc modifications are made to overcome problems such as those posed by the need to transport ecological communities over large distances and subsequently deposit them mostly intact.
Sedimentary, my dear Watson
Flood geologists have recognized several other problems for the EZ model. For example, one would expect serious erosion and transport of sediment in the worldwide flood. Acknowledging that most present-day sediment cover is found on the cratons of the continents and that the surrounding ocean basins have very little sediment contained within them, Kurt Wise and others (1994) concede that, if the Flood had occurred, the continental rock should have been eroded and subsequently deposited as sediments in the ocean basins — which is clearly not the case. They are forced to postulate that all the sediments and rocks that now cover the continents came from the area of the ocean basins and continental shelves. The mechanism they propose is complex and beyond the scope of this article, but clearly the creationist model requires that virtually all of the sediment that now is contained in sedimentary rock on all the continents was transported over considerable distances to its present location. Taken in conjunction with the earlier observations about geologic deposits in the North American epeiric seas, this means that all the sediments, as well as most of the terrestrial flora and fauna, must have been transported over great distances before being redeposited in an arrangement that resembles their original ecological relationships — and all within a very short time. And time presents the next problem for the flood model.
Readers of RNCSE know that young-earth creationists (YECs) are fond of arguing that the many types of isotopic age determination are based on a series of false premises, untenable assumptions, and biased calculations (see Dalrymple 2000; Thomas 2000; York and Dalrymple 2000). However, there is one aspect of geochronology that is incontrovertible even by such creationists: that isotopic age determination yields progressively older dates for progressively lower stratigraphic levels — even though YECs refuse to accept that these methods accurately estimate the ages of these deposits in the millions of years. This consistency among methods for producing older ages in lower strata is thoroughly verified (Dalrymple 1991). From the geological literature, it is clear that the ages of the upper and lower strata are not separated by a few months (as the flood model requires), but by millions of years.
Since this determination of older ages in lower strata is based on a different relative proportion of isotopes of radioactive elements, it is fair to ask how the flood model accounts for this situation. In essence, the flood model requires that the sediment and its component radioactive elements, after being transported quickly and forcefully over great distances, came to rest in a manner that would place radioactive materials that yield the oldest dates in the lowest strata with progressively higher strata showing younger dates by virtue of a smaller proportion of isotopic decay products. There is no known mechanism that could allow for such segregation. After all, hydrodynamic sorting is based on the qualities of density, buoyancy, and displacement, not on isotopic composition, which should have no effect on the placement of these deposits.
Organic change correlates with geologic time
Another significant feature of the stratigraphic record that cannot be explained by transport is the clear "vector" of biological complexity that runs throughout the geologic column. In recent years some creationists have claimed that organic complexity does not increase from lower (older) to higher (younger) stratigraphic intervals. Although there was never any doubt among paleobiologists about the falsity of this claim, it has, nevertheless, been decisively answered by James Valentine and others (1994) in an article in Paleobiology entitled "Morphological complexity increase in metazoans". Valentine and his coworkers demonstrated that organic complexity indeed increases afrom lower-to-higher stratigraphic intervals.
Taking somatic cell types as their comparative measure of complexity, Valentine and his coworkers noted that at the older end of the scale, the placozoans (which have no tissues, organs, organ systems, heads, or tails) possess only four somatic cell types, while mammals, emerging much later in evolutionary history (represented in their study by hominids), possess over 200 somatic cell types. Plotting times of origin of body plans against cell type numbers, they discovered that complexity, as measured by the number of somatic cell types, increase in the fossil record, at an average rate of about 1 cell type per 3 million years.
Clearly, organic complexity does increase throughout geologic time.
Of course, this finding completely contradicts any model according to which the pattern of deposition of fossil organisms was produced by their transportation in a single violent flood into their present positions. What mechanism can flood geologists invoke to explain how organisms naturally subject to simple hydrodynamic principles nevertheless should end up in the fossil record in a manner that mimics the pattern that we would expect to find if organic evolution were true?
Tracks and traces
A further problem for the flood model is the abundance of trace fossils and tracks: there is no significant stratum in the Mesozoic (or even later in the Cenozoic) in which we cannot find either individual prints (ichnites) produced by terrestrial organisms or plentiful terrestrially-produced suites of trace fossils created by ecologically related organisms (ichnocoenoses). This means that at no time in the Mesozoic was the earth so completely flooded that terrestrial organisms could not walk upon the surface. If, in fact, a global flood had covered the earth during the Mesozoic, terrestrial animals hardly could create trace fossils on the surface! Thus at no time during the Mesozoic was the earth entirely covered by water.
The implications of fundamental geologic and paleontologic observations for flood geology are profound. When creationists are forced to realize that the geologic column cannot be explained in terms of transport mechanisms, then they are forced to recognize that the geologic column represents the product of natural forces acting over an immense span of time. A corollary of this recognition is that measurable morphological change, so richly documented by the fossil record, is real and indeed, a product of evolution. However, the response typical of YECs is to invent ad hoc modifications of their models in an attempt to make observations consistent with a short-term worldwide flood. In every case, the standard evolutionary and geological models produce satisfactory explanations of the observation that are corroborated by independent research in several scientific fields.