What, then, is cladistics? Cladistics, or phylogenetic systematics, is a way of analyzing relationships that was first brought to the fore in the late 1960s, although it had been proposed in Germany decades earlier. By the early 1980s, it had demonstrated its practical and theoretical value to enough of the community of systematists that its methods became commonplace in studies of all branches of organisms, in most top scientific journals, and in the National Science Foundation's decisions about awards in systematic biology. Its influence has grown in succeeding years to the point that statements about evolutionary relationships are no longer taken seriously in the community of systematists unless backed by a cladistic analysis. This is true regardless of the type of organism and regardless of whether the postulated relationship is based on morphology, molecules, behavior, or fossils.
There is no guarantee that any given cladogram will not be revised or overturned by further study (new techniques are constantly being developed and revised); they are hypotheses that are meant to be tested, after all. But cladograms, unlike any other kind of evolutionary hypothesis of relationships, are explicit in their methods and the data on which they are based, and are testable. This gives them practical value. And because they restrict evidence to new, unique evolutionary features as a way of determining relationships among closest relatives, they are more consistent theoretically with the expectations of evolution than any other method.
Cladistics and its Critics
Critics of cladistics (those who still remain), or critics of the dinosaur-bird hypothesis, claim that cladistics has become dogma. To understand how people in the field respond to this complaint, I suggest the following analogy.Most people accept, on practical as well as theoretical grounds, that medical imaging (X-rays, CAT scans, and so on) shows what is going on inside a patient. Before X-rays, physicians had to operate (dangerous and painful) or feel around on the outside and infer the patient's condition. With modern medical imaging, physicians can easily see an intestinal blockage, a tumor, or a fracture. The people who object to cladistics, decades after its general adoption, are like doctors who would rather feel around to diagnose the problem. It is not that they are necessarily wrong, but now we have better ways to diagnose ailments. If you were sick, would you rather have your doctor just feel around, or use an imaging technique such as an X-ray? At least as a second opinion?
Another criticism, focused on the widespread use of computers in cladistic analysis, is that cladistics is just "garbage in, garbage out". Not so: the computer does not do the thinking for the scientist. The scientist determines which characters and organisms to choose and which states are primitive and derived. The computer's role is just to do electronically what takes much longer to do by hand: namely, sorting out the shortest and simplest evolutionary "family" trees for further testing. It is exactly analogous to using a calculator instead of pencil and paper to add a long list of figures. And every cladistic analysis contains a list of the characters and organisms used and how the character states were coded, so anyone can run the analysis again - with variations, if desired.
Why the emphasis on methods in an article that is supposed to be an update on the dinosaur-bird hypothesis? Because every couple of months - or so it seems - there is some kind of challenge to the hypothesis, mounted by the same cast of characters. Well, fine; science is built on challenges to what we think we know. But when do we start to decide that a hypothesis is pretty robust to all this testing, and what standards of testing should we require? Although it has been over 25 years since Ostrom put forth the dinosaur-bird hypothesis, its opponents have yet to propose an alternative, testable hypothesis. So far not a single one of these opponents has ever - and I doubt they ever will - come out and said, "here's another animal or group of animals that we propose as closer to birds than the theropod dinosaurs, and here are the reasons." Their hypothesis is simply that the dinosaur-bird hypothesis is wrong. All the proposed similarities of birds and dinosaurs are mistakes and delusions.
Opponents also claim that the dinosaur-bird hypothesis is dogma, apparently on the grounds that those who accept it have not accepted the opponents' arguments for rejecting it. But science does not require unanimity, it does not force agreement, and it does not settle issues by vote. Some geologists went to their graves not accepting that the continents move. Science progresses nonetheless, by the accumulation of evidence and the testing of hypotheses that account for it. Today it is difficult to find an article in geology that begins by allowing that plate tectonics is only one possible model among many other equally plausible ones - even though 40 years ago the theory was hotly contested.
Well, what methods and tests are the anti-theropod critics using? Not cladistics: they do not use cladistics, because every time someone does a cladistic analysis, birds come out most closely related to theropod dinosaurs. The critics often admit their aversion to cladistics, but even when they do not, their papers speak for them: not a single real cladogram has appeared in any of their works.
Okay, we can all agree that any hegemony of method can be challenged. But in science, we do need methods. What, then, do they propose in place of cladistics? The answer is a resounding silence. They will not say what methods they are using, and so it is impossible for anyone to test their statements. Occasionally, they claim that they do not need methods because they have the crucial evidence to falsify the dinosaur-bird hypothesis. Luis Chiappe and I dealt with these objections in several publications, including our 1998 article in Scientific American (Padian and Chiappe 1998a) and a longer, more technical one in Biological Reviews in the same year (Padian and Chiappe 1998b).
There are two larger points of interest here. I am often asked, by other scientists, by reporters, and by members of the public who are just interested in questions about dinosaurs and evolution, "So what is it with these anti-theropod people? It sounds like you are arguing with creationists." And here, especially for the NCSE audience, I would like to demur on this comparison. It is intellectually dissonant to mention these two groups in the same sentence, because obviously the dissenters to the dinosaur-bird hypothesis are competent scientists who accept evolution. But the comparison appears to recur because if you have no alternative hypothesis to test scientifically, and you do not accept the methods of the field yet have no alternative methods that can be used, at some point observers will begin to wonder about the scientific basis for your statements. I think, in fairness to these dissenters, that they hold that evolutionary processes, as they understand them, would not be able to produce birds from dinosaurs; so the evolutionary patterns that we see in cladograms must be wrong.
The second point has to do with public education. Why does the public not understand the methodological basis of this dispute? The answer is two-pronged. First, the reporters assigned to cover the story do not make the issue clear to the public. This is because most of them do not understand it; they think that the dispute is largely motivated by personalities and politics in the absence of definitive evidence. It could be improved if reporters would explain at least a bit about the methods and standards of evidence, as opposed to "he said, she said" journalism. But after all, they are journalists, and we cannot expect them to be scientists too, any more than scientists can be competent reporters.
The second reason, which encompasses the first, is that even though cladistic analysis has been the standard for the field for two decades, it is almost unknown to the general public. Textbooks continue to teach the outworn Linnaean system and to portray taxonomy as some kind of art, instead of as a process that arranges organisms according to scientifically tested hypotheses about the evolutionary changes that produced a variety of descendants from a common ancestor.
Recent Developments in Bird Evolution
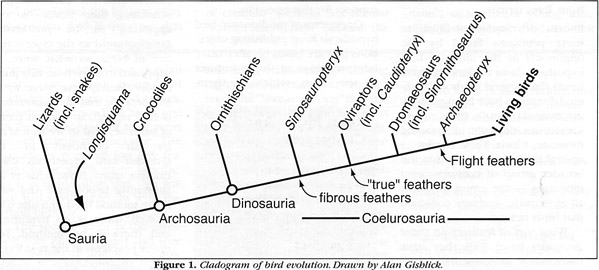
Recent developments illustrate some problems faced by the opponents of the dinosaur-bird hypothesis. Most readers will have heard that several kinds of feathered dinosaurs have been recently discovered in the Early Cretaceous deposits of Liaoning Province, China. They belong to several distinct groups within the broader group of coelurosaurian dinosaurs - the group to which all systematic analyses conclude that birds belong.What sort of feathers do these dinosaurs have? Well, they have two kinds of integumentary structures. One kind produces a thick, relatively short and dense pelage made up of fibrous, filamentous structures that appear all over the body. These, claim some opponents of the dinosaur-bird hypothesis, are merely collagen (a common connective tissue in skin). However, molecular analysis shows that these structures are made of keratin - and not just any keratin, but the beta-keratin that feathers have (and, equally important, not the alpha-keratin that make up the scales of today's reptiles). Dinosaurs that have this kind of integumentary structure include several coelurosaurian theropods, such as the compsognathid Sinosauropteryx, the therizinosaurid Beipiaosaurus, and the dromaeosaur Saurornithosaurus (listed in order of their closeness to birds). More feathered coelurosaurs continue to be discovered and described in the scientific literature.
The second kind of integument is true feathers, which have a central shaft, two vanes, and barbs. These true feathers are attached to the forelimbs and tail just as the feathers of Archaeopteryx and living birds are. True feathers are found in the oviraptorosaur Caudipteryx and another form, Protarchaeopteryx, which are coelurosaurs. Opponents of the dinosaur-bird hypothesis have claimed that these are merely birds that have given up the ability to fly, but because they have not performed a phylogenetic analysis of any sort, they have no support for this assertion.
Opponents of the dinosaur-bird hypothesis keep publishing objections that are based on alternative interpretations of single features or specimens, which by themselves do not falsify the dinosaur-bird hypothesis. The most recent is a re-interpretation of Longisquama, an enigmatic reptile of undetermined relationship that occurs in the Triassic of Kazakhstan. It has been known for 30 years, but what is most interesting about Longisquama are the long oblong structures that appear to emanate from its vertebral column. Each of these structures has a central stalk that separates two flat, semi-elliptical surfaces. There are no barbs, but there are some features emanating from the stalk, wavy in contour, directed proximally near the base of the stalk and distally near its end. The entire structure is surrounded by a perimeter reminiscent of a rubber band.
The re-interpretation of this specimen as possessing true feathers (Jones and others 2000) was supposed to overturn the dinosaur-bird hypothesis, according to an aggressive press release and statements made for the benefit of the media, but for the most part it just left paleontologists scratching their heads. No one appeared to be postulating Longisquama as the closest relative of birds, so what were we supposed to learn from this publication? Perhaps we were to be admonished that the fossil record is rich enough to contain plenty of surprises, and so we should not be so confident in the dinosaur-bird hypothesis. Okay, caution taken. Now, what is the alternative hypothesis? And what is the method used to frame it?
Well, there is no hypothesis, and there is no method. Two major problems in the re-interpretation of Longisquama indicate the pitfalls of the "alternative" approach. First, the opponents of the dinosaur-bird hypothesis who published this paper asserted that Longisquama was an archosaur, but it is not. Archosaurs (by definition) include birds and crocodiles and all descendants of their closest common ancestor. No analysis yet has placed Longisquama anywhere near this group. Rather, it is apparently somewhere within Sauria, the broad group that includes living lizards and snakes, Sphenodon, crocodiles, birds, and all the descendants of their most recent common ancestor. The specimens preserve too few features to be much more specific. So it is difficult for them to propose that this animal had anything to do with the origin of birds.
The second error is the assertion that Longisquama had true feathers (Reisz and Sues 2000). Reporters found it difficult to get anyone else to agree with this (Stokstad 2000). The two most noted experts on feather structure and development rejected the idea, and one opined that the paper would not have been published in even a third-rate ornithological journal. As noted above, the similarities to feathers are superficial at best. Why, then, did the paper receive such attention in the popular and scientific press? Well, scientific journalism, especially in high-profile journals, is not above a bit of the "Man Bites Dog" mentality; there is competition to report on what seems new and exciting, even in the news sections of peer-reviewed publications.
Let me propose a litmus test. Next time you encounter a newspaper or television story on this or any scientific issue, get to the bottom of it with two questions: (1) What exact hypothesis is being proposed here to supplant another one (and it cannot be simply that the first hypothesis is wrong: we assume that in all tests)? (2) What methods are being used, if not the standard methods in the field, and how do we know that these are better than the standard methods? If and when the opponents of the dinosaur-bird hypothesis manage to give satisfactory answers to these two questions, they will be taken seriously.